It is estimated that 50-60% of all conceptions in normal women abort spontaneously before pregnancy is even detected. Another 10-20% abort later in pregnancy. Chromosomal abnormalities appear to cause about 40-50% of spontaneous abortions. Some chromosomal abnormalities are tolerated and result in a variety of developmental disorders, most familiar is Down's syndrome and retinoblastoma. Many chromosomal anomalies arise during gametogenesis, fertilization and cleavage as a result of abnormal meiosis or mitosis: nondisjunction, translocations, deletions, or rearranged segments. The absence of a specific chromosome in a gamete that combines with a normal gamete to form a zygote results in a condition known as monosomy. The presence of two of the same kind of chromosome in one of the gametes that forms a zygote results in trisomy
Down's syndrome is an example of a disorder caused by an error during meiosis or mitosis. If the two copies of chromosome 21 fail to separate during the first or second meiotic anaphase of gametogenesis in either parent (nondisjunction) half the resulting gametes will lack chromosome 21 altogether and the other half with have two copies.
Monosomy 21----embryos rapidly die
Trisomy 21---down's syndrome
What happens if chromosome 21 nondisjunction occurs in a single embryonic cell during cleavage? Mosaic of normal and trisomy 21 cells. The severity will depend on when and where the chromosomal defect occurs during development.
2010 Nobel Prize in Medicine to Robert Edwards for the development of In Vitro Fertilization (link to lecture honoring him) (IVF technique REQUIRED)
FERTILIZATION
(Historical review)
Scientists tend to look for the best biological system to study a particular problem. Our strong belief in evolution and the unity of all life convinces us that much of what we learn in one animal will illuminate our understanding in all animals. Comparisons of developmental processes in different animals allows us to decide what features of development are most fundamental and what problems or mechanisms are species specific.
Most of what we know about fertilization was first discovered in sea urchins. It is relatively easy to get thousands of gametes and fertilization takes place externally in seawater. It is much more easily studied than internal fertilization. As you read in Gilbert, it was Oscar Hertwig who chose the sea urchin as a good system to examine fertilization. Because the sea urchin egg is very transparent he could watch the entire process of fertilization under the microscope. He clearly described the entry of a single sperm into the egg and the fusion of the sperm and egg pronuclei to form a single nucleus from which all subsequent nuclei were derived.
Fertilization can be divided into four main parts
1. Recognition events between sperm and egg: species specificity
2. Regulation of sperm entry into egg: block to polyspermy
3. Fusion of sperm and egg genetic material
4. Activation of developmental process within the egg
Structure of the gametes
Sperm
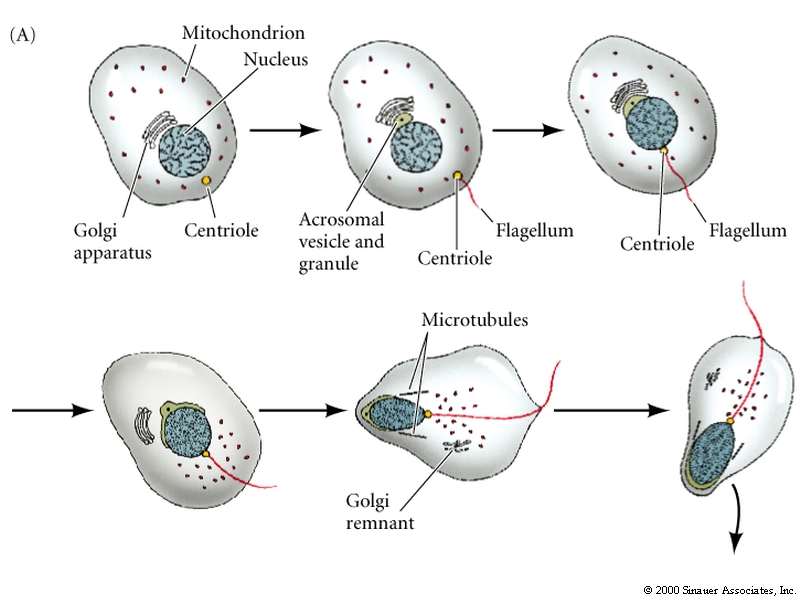
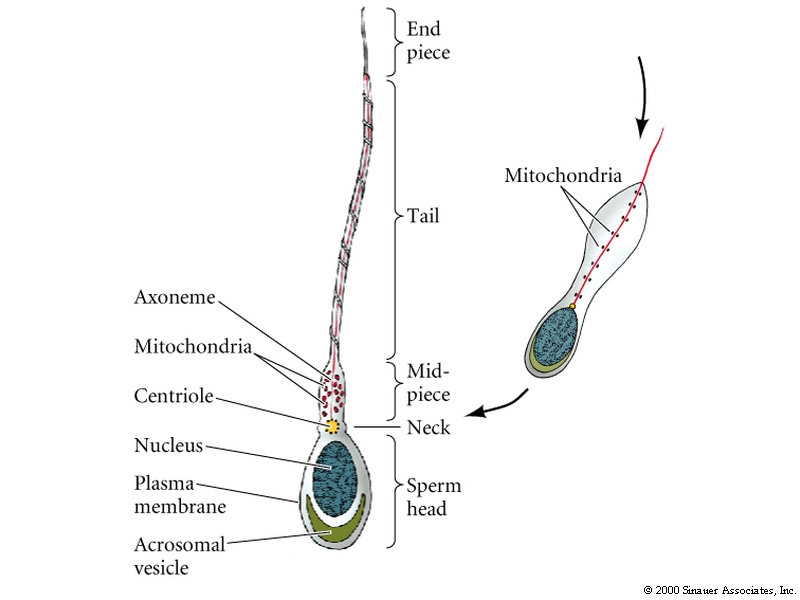
Haploid nucleus, flagellum, and specialized molecules to recognize and penetrate the egg.
DNA must undergo a remarkable condensation to fit into the sperm head. It requires a special set of packing proteins, called protamines (32aa, very arg rich), that replace the histones proteins and cause the DNA to pack into an almost crystalin form.
The acrosomal vesicle is basically a specialized lysosome derived from the golgi. A store of bound actin monomer between the nucleus and the acrosomal vesicle drives the extention of the acrosomal process by a process of rapid polymerization.
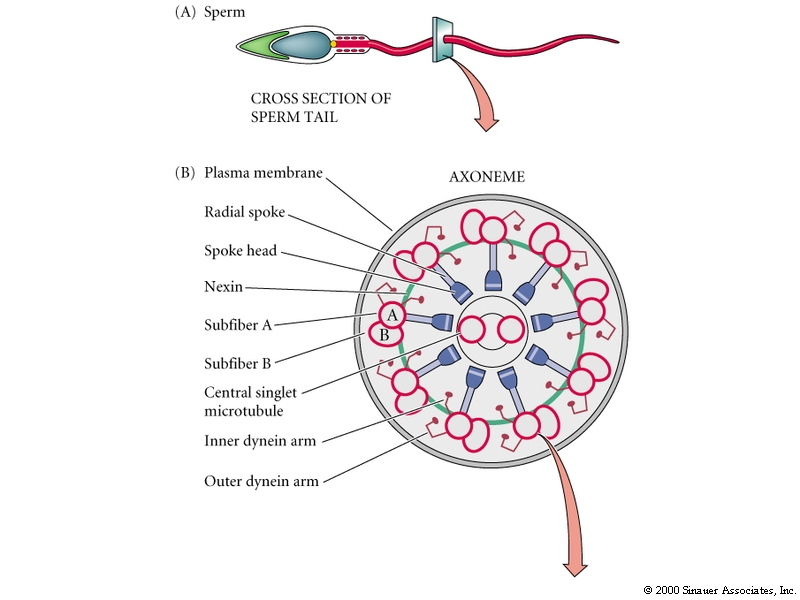
Sperm are propelled by an axoneme motor made up of specialized microtubules and dynein, an ATPase that converts the hydrolysis of ATP into mechanical energy. See the link for a sperm animation http://www.luc.edu/faculty/wwasser/dev/devm.htm.
Egg
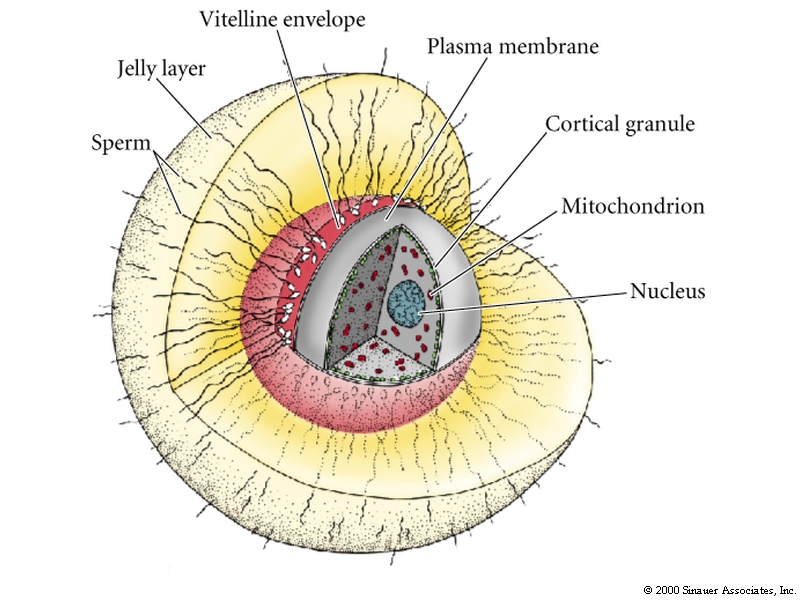
The egg is rich in protein, rRNA, mRNA, "morphogenetic factors" and of course more or less yolk protein for maintenance of the developing embryo.
Haploid nucleus
Yolky cytoplasm
Mitochondria (only maternal survive)
Cortical granules
Modified endoplasmic reticulum like compartment for sequestering Ca
Plasma membrane
Vitelline membrane (mammalian zona pellucida)
Jelly layer (sometimes contains chemoattactant for sperm)
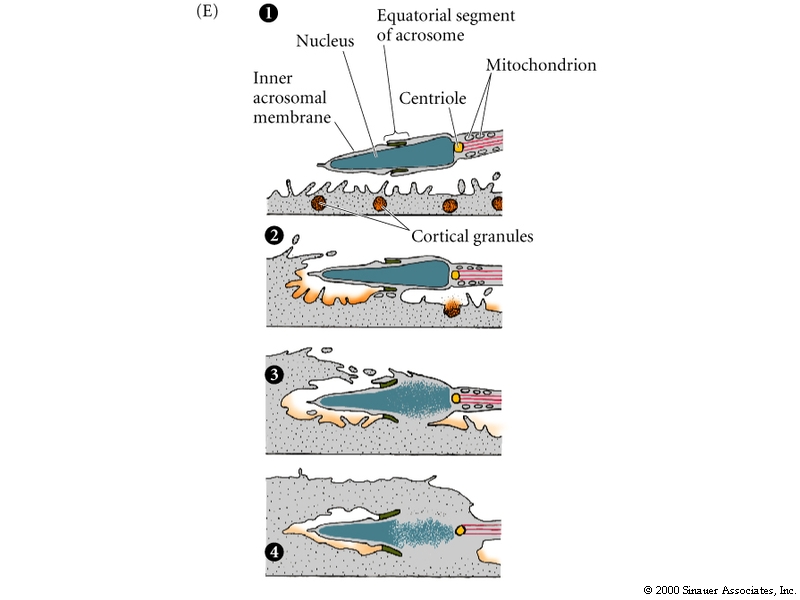
Contact with the Jelly Coat of an Egg Stimulates a Sea Urchin Sperm to undergo an Acrosomal Reaction
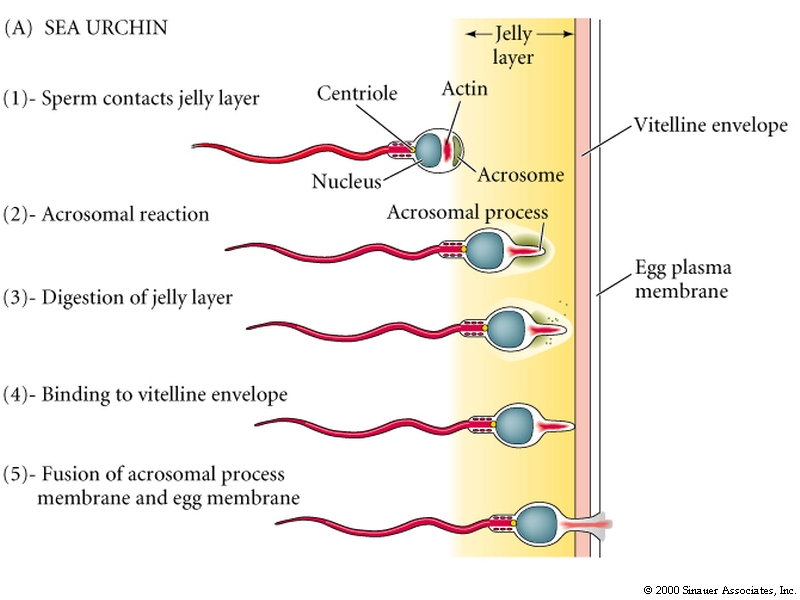
The contents of the acrosomal vesicle are released and acrosomal process extends. The acrosomal process is covered by the acrosomal vesicle membrane and some of its contents.
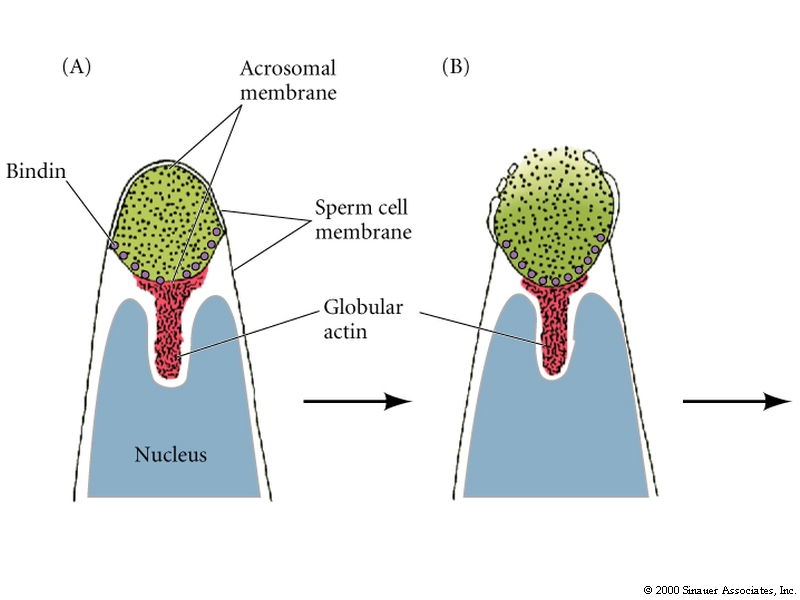
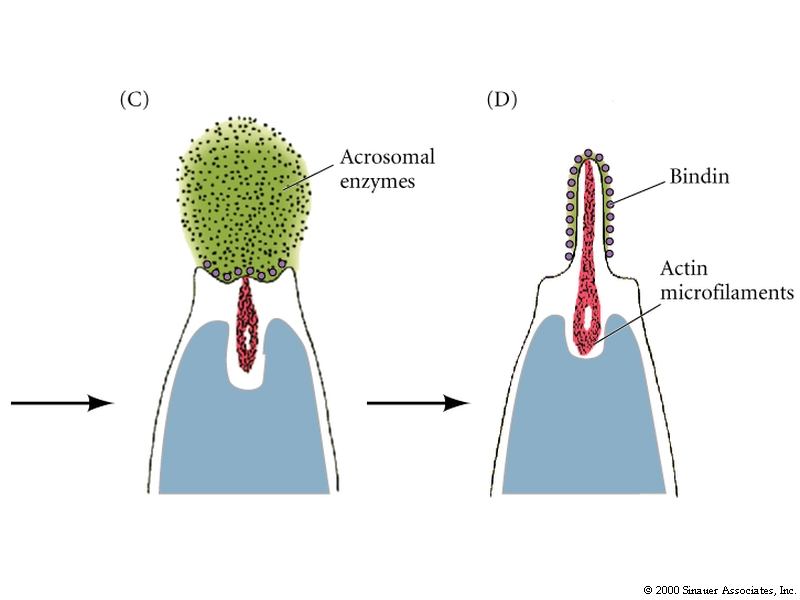
Acrosomal vesicle contains:
1.Hydolytic enzymes that dissolves molecules in jelly coat.
2. Specific binding proteins that mediate attachment between sperm and egg vitellin membrane(Bindin in Sea Urchin).
3. Hydrolytic enzymes to break down the vitellin membrane so the sperm can reach the egg plasma membrane.
The acrosomal reaction in (in sea urchin) is triggered by a fucose sulfate rich polysaccharide in the jelly coat. This polysaccharide binds to a receptor molecule in the sperm plasma membrane that causes the sperm cell to depolarized by Na. This induces voltage gated Ca channels to open and a proton/Na exchange pump to increase intracellular pH.
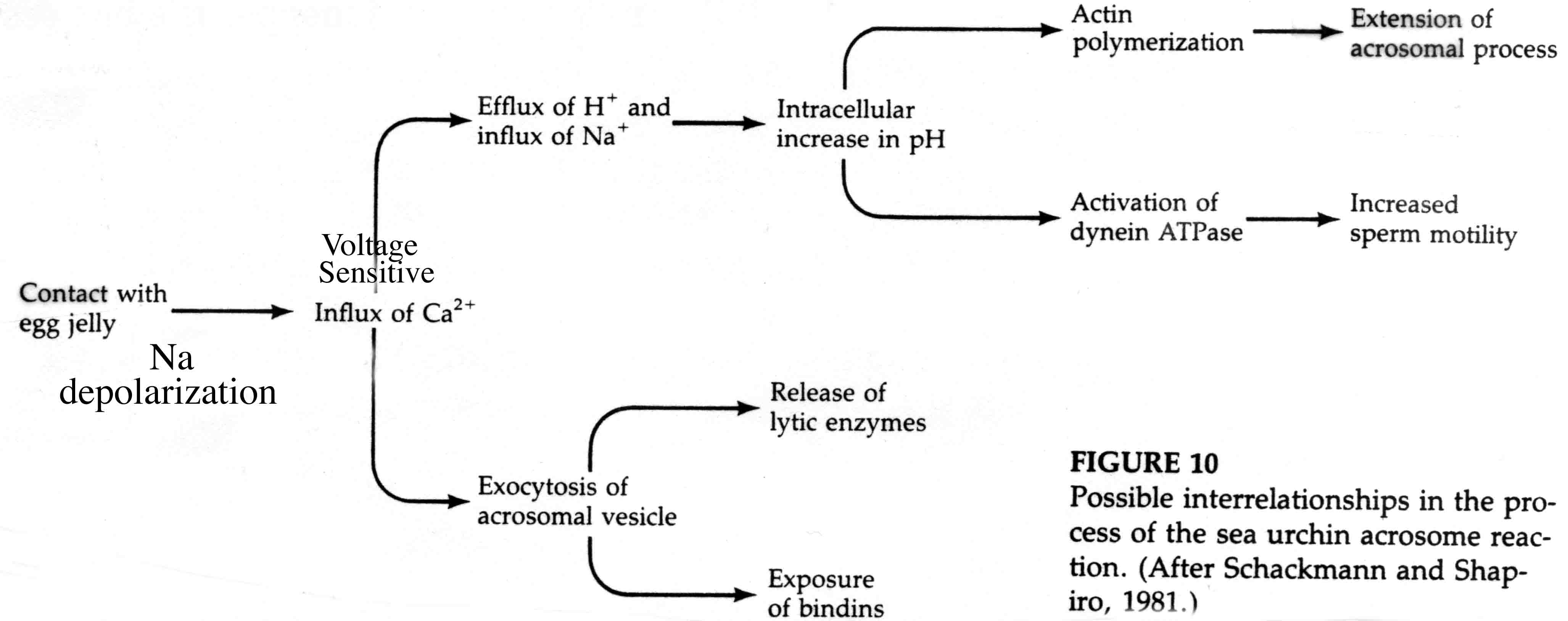
The increase in intracellular pH and Ca initiates the acrosomal reaction. Ca is necessary for membrane fusion and the increase in pH is thought to reduce the binding affinity of an actin binding protein that inhibits actin polymerization. This leads to the explosive polymerization of actin to form the acrosomal process.
The increase in intracellular pH is also responsible for activation of dynein ATPase and a subsequent increase in sperm motility.
In the mouse the process if very similar, but note the specific differences. Sperm penetrates cumulus layer instead of jelly layer and acrosomal reaction is stimulated by contact with Zona pellucida.
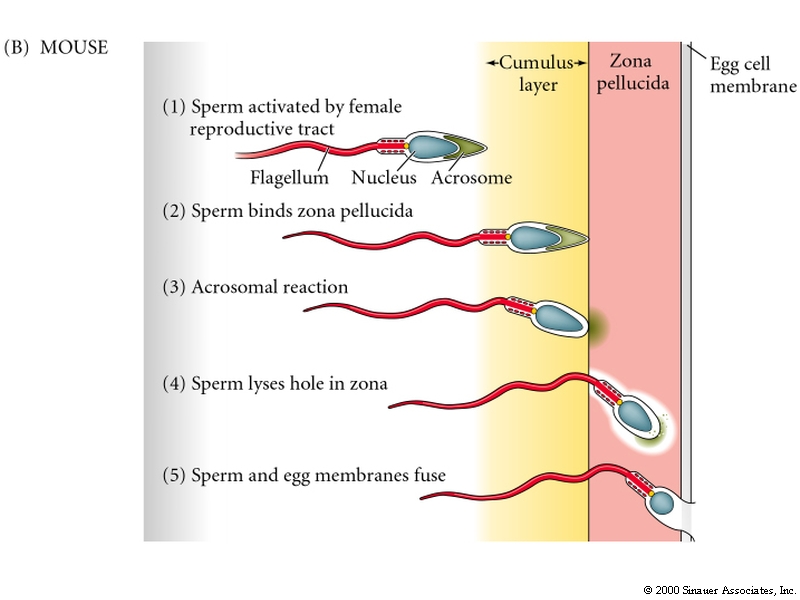
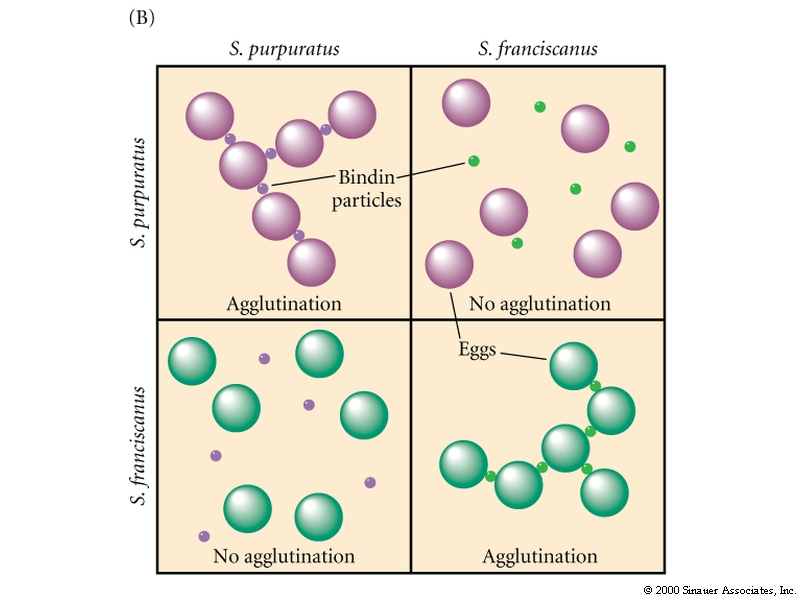
Sperm Egg Adhesion is mediated by Species specific macromolecules
A protein call BINDIN is sequestered in the acrosomal vesicle and after the acrosomal reaction coats the surface of the acrosomal process. The receptor for BINDIN is a proteoglycan located in the vitelline membrane. It is species specific. Specificity seems to reside in the sugar moieties. Bindin acts as a lectin. In addition BINDIN seems to have a membrane fusion activity when tested in vitro. It may be the molecule responsible for fusion of the sperm and egg plasma membranes. The experiment shown below illustrates the species specific adhesion activity of the molecuele Bindin.
In the mouse the sperm-egg binding activity is mediated in part by ZP3.
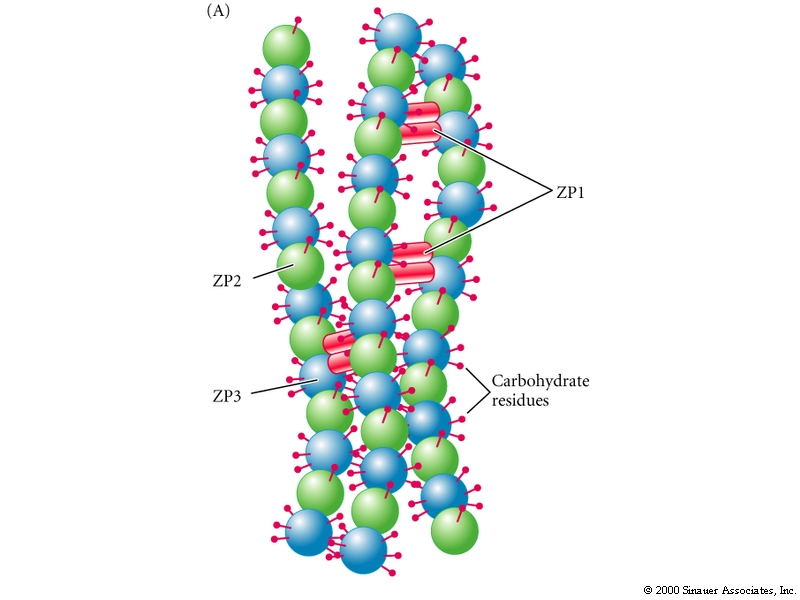
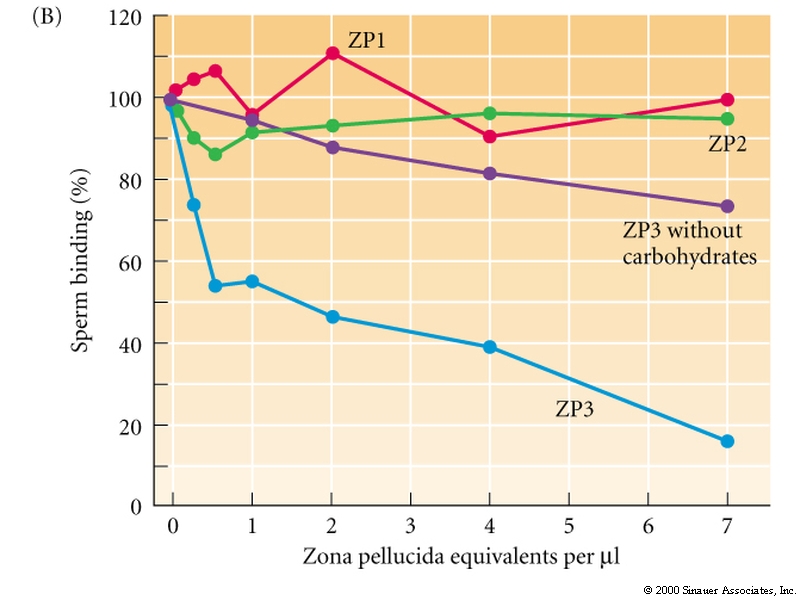
The above figure demonstrates the role of ZP3 in sperm binding to the Zona pellucida. The carbohydate seems particularly important for binding .
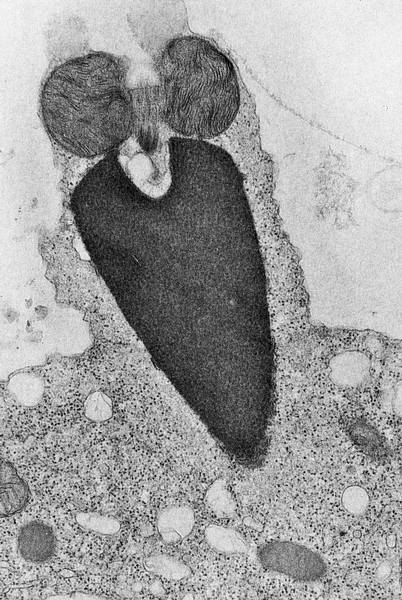
This transmission electron micrograph from your text shows the sperm membrane fused with the egg cell plasma membrane and the entry of the sperm nucleus into the egg cytoplasm.
Egg Activation involves changes in intracellular ion concentrations
The unfertilized egg is metabolically dormant--it does not synthesize DNA and only small amounts of RNA and protein. Activation by sperm leads to DNA synthesis and cleavage. HOWEVER, the program of egg activation does not require sperm, the sperm is simply a signal that initiates the program. Eggs can be activated by many nonspecific treatments, such as a needle prick. Egg activation also does not require any new protein synthesis (cyclohexamide does not block).
Sperm Activation of the egg causes:
1. Increased Na current leading to depolarization within 1/10 of a second.
2. Release of Ca from intracellular stores that raises Ca concentration within 10 secs.
3. Coupled influx of Na and efflux of H leads to pH increase in first 60 sec
The rapid depolarization of the egg plasma membrane prevents further sperm egg fusions----This is the fast block to polyspermy.

In most animals that have external fertilization the sperm can fertilize the egg at many places on its surface. Here a rapid depolarization (-60 to +20mv) caused by the binding of the first sperm prevents other sperm from fusing with the egg plasma membrane for about 1 minute. The membrane potential then returns to normal. You can prove that its membrane potential alone that is responsible by artificially depolarizing an egg with a microelectrode and demonstrating that sperm can bind to the egg, but cannot fuse for as long as you keep the egg depolarized. Implies that there is a voltage sensitive ligand whose conformation is changed with depolarization and prevents fusion.
If more than one sperm fuses with the egg extra mitotic spindles form, chromosomes don’t assort correctly and development is arrested.
Some animals such as fish and insects have a small channel, call the micropyle, though which the sperm have to pass to reach the egg plasma membrane. Only a single sperm at a time can pass through this channel and after the first sperm pass through it stimulates the release of the egg cortical granules which blocks the passage of subsequent sperm.
The cortical reaction is responsible for the slow block to polyspermy
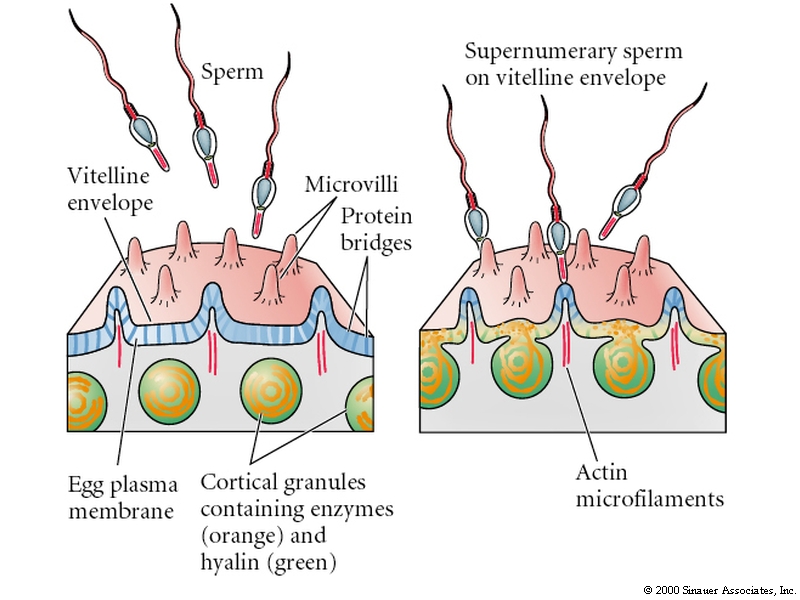
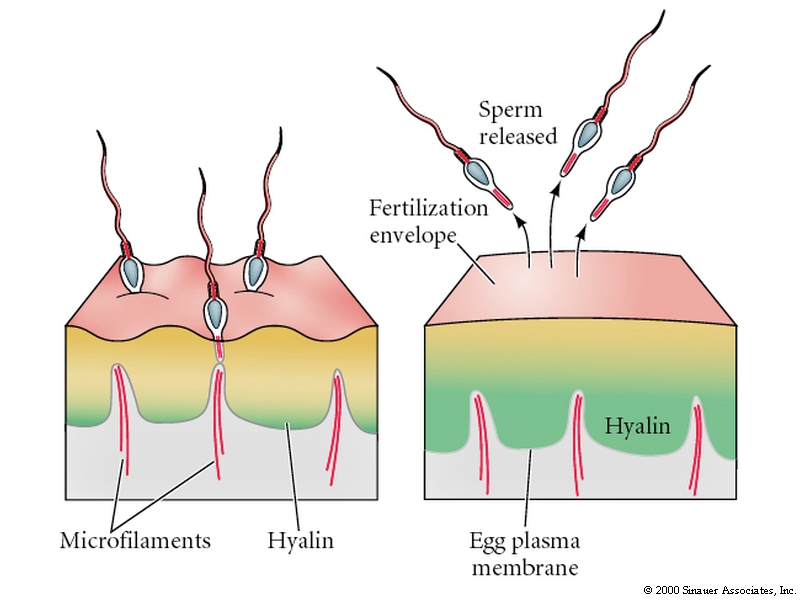
The increased Ca concentration causes the cortical granules (about 15000 at 1 um) to fuse with the egg plasma membrane and release their contents. The enzymes released by the cortical granules inactivates the BINDIN receptors on the vitelline layer and a peroxidase crosslinks tyrosine residues of adjacent proteins within the vitelline layer so that sperm can no longer attach to or penetrate to the egg. Additionally, attachments between the vitelline layer and the egg are digested releasing many osmotically active particles into the space between the vitelline and plasma membranes. Water rushes in and forces the vitelline layer away from the surface of the egg. It is now termed the FERTILIZATION ENVELOPE. Here is a movie by Rachel Fink and the Society for Developmental Biologists from "A Dozen Eggs".
HYALIN from the cortical granules forms a protective and supportive protein coat around the fertilized egg (now embryo).
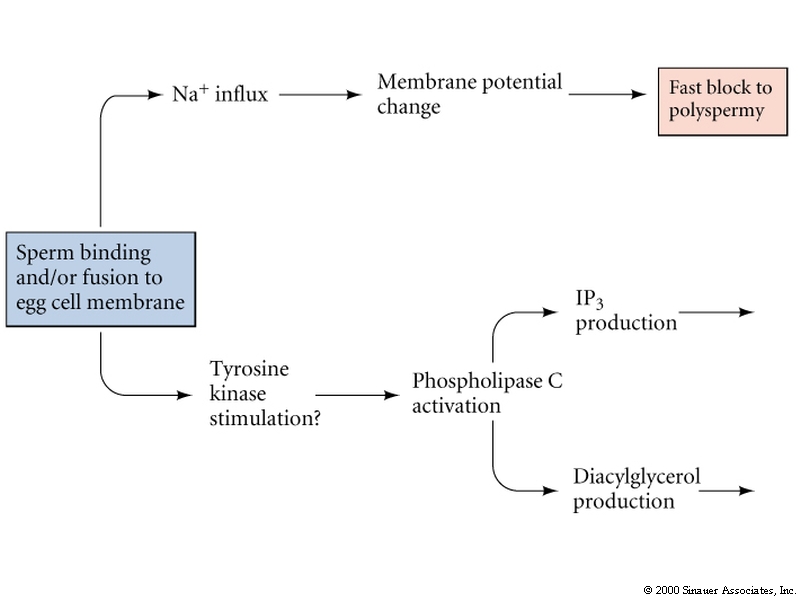
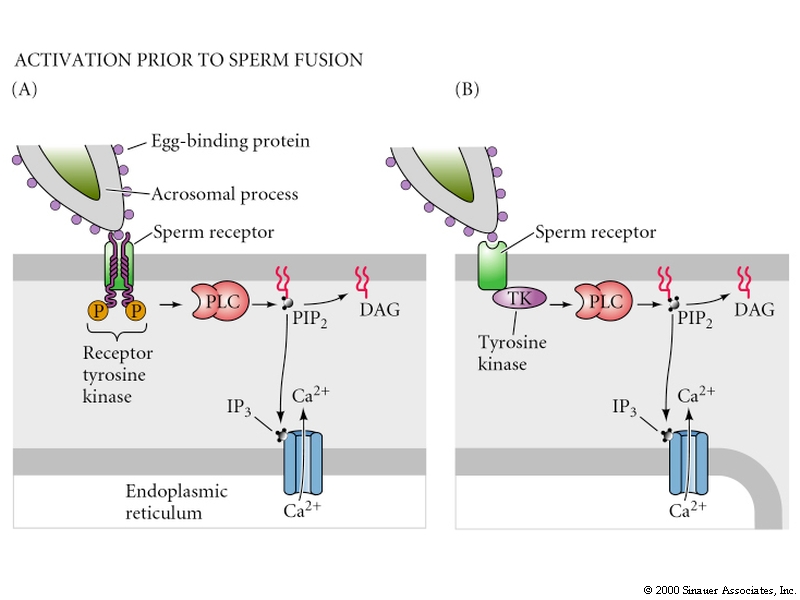
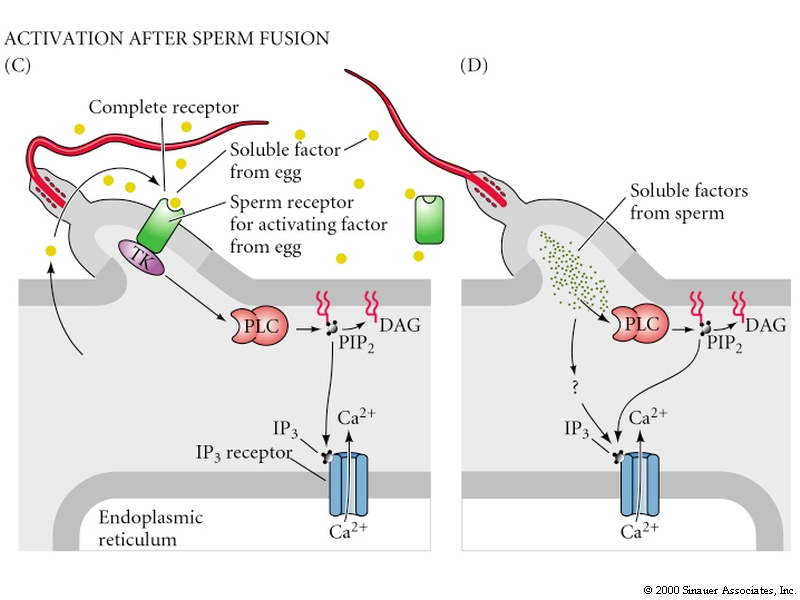
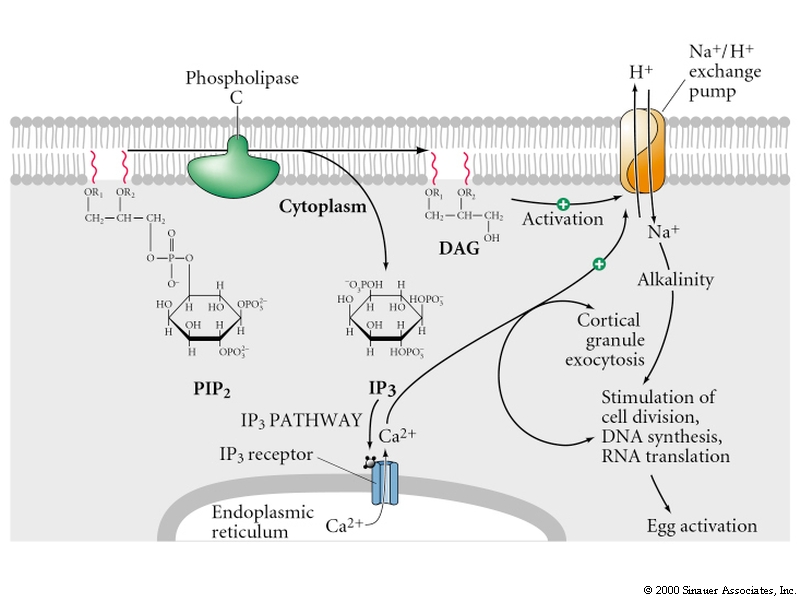
Egg activation is mediated by the inositol phospholipid cell signaling pathway.
Remember, the early change in membrane potential is only necessary for the fast block to polyspermy. Egg activation is initiated by the rise in Ca concentration. Sperm cell ligands bind to egg membrane receptor proteins that either directly or indirectly activates a tyrosine kinase which in turn activates phosphoinositide specific phospholipase C and leads to hydrolysis of phosphatidylinositol bisphosphate (PIP2) in the plasma membrane to produce inositol trisphosphate (InsP3) and diacylglycerol. The InsP3 causes release of Ca from an internal store (specialized endoplasmic reticulum).
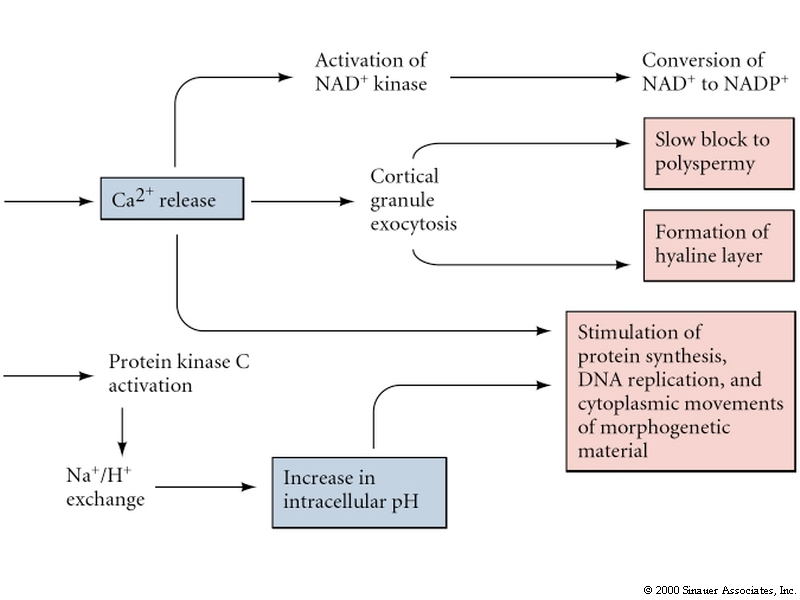
Protein Kinase C is activated by diacylglycerol and Ca. It phosphorylates specific serine and threonine residues on target proteins. In this case a Na/proton exchanger which causes the intracellular pH to increase from 6.7-7.2. The low pH is thought to keep the egg metabolically inactive. Initial protein synthesis is due to translation of maternal mRNAs and maternal rRNA. The increase in pH is thought to make the mRNAs available for translation and increase the efficiency with which rRNA move along the mRNA. DNA synthesis is also facilitated. Rise in intracellular pH in some organisms induces the late synthetic events of egg activation
The following figures show possible alternative activation pathways in cartoon form.
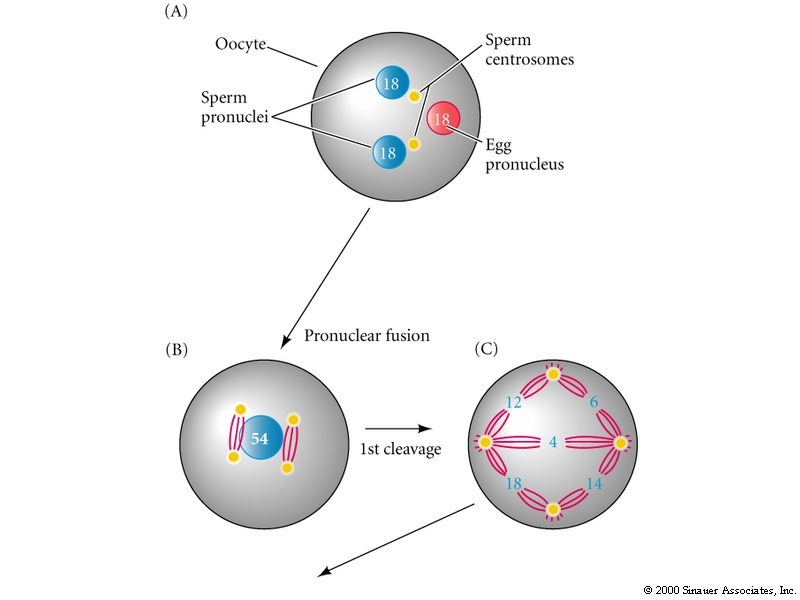
Fusion of sea urchin sperm and egg pronuclei depends on centrioles donated by the sperm
Sea urchin sperm donates not only the haploid nuclei, but also two centrioles. These centrioles form the sperm aster (microtubule array) which is necessary for guiding the male pronucleus to the female pronucleus. On contact the nuclear membranes of the two pronuclei fuse to form the diploid nucleus of the zygote. The pair of sperm centrioles divide with their associated asters and form the two poles of the first mitotic spindle in preparation of the first cleavage division.
In contrast, during mammalian fertilization it's the egg that contributes the centrioles, and the male and female pronuclei do not fuse till the nuclear membranes dissolve in preparation of first cleavage.
Recently scientists have been able to generate eggs and sperm from ES cells. They have used these "made" sperm and eggs to produce normal mouse pups! Neither eggs from females or sperm from males is necessary to produce a normal embryo. Here is a link to article that will work from off compus sites.
New research shows that time lapse imaging of early cleavage stage blastomeres can predict defective embryos that will spontaneously abort and therefore should not be used for implantation into a womens uterus. Should advance success of IVF technique. (Paper Required)
Germ line therapy can potentially cure maternally transmitted mitochondrial disease and allow women to have healthy children. Online and local link.
NIH Mitochondrial diseases Page